
Hipoteza hybrydowa
3: Analiza wstępna
https://www.macroevolution.net/hybrid-hypothesis-section-1.html
|
Wtedy poczułem się jak obserwator nieba,
Kiedy nowa planeta wpływa w jego pole widzenia. — John Keats
|
(Ciąg dalszy z poprzedniej strony )
|
Bardzo podobał mi się ten fragment o pochodzeniu człowieka. Wyraźnie dobrze zbadany, starannie opracowany i bardzo przekonująco napisany. Podzieliłem się wieloma jego bardziej uderzającymi punktami z moją dziewczyną, która jest Chinką (obecnie jestem w Chinach) i której dieta zawiera dużo wieprzowiny, jak to jest w Chinach zwyczajowe. Mówi, że zaczyna czuć się naprawdę źle z powodu jedzenia świń.
— Chase Dumont
|
Podczas gdy niektóre świnie domowe są hodowane tak, aby były stosunkowo bezwłose, wszystkie dzikie świnie wydają się być w pełni owłosione. W rzeczywistości, gdy świnie domowe stają się dzikie, natychmiast powracają do owłosionej formy. Jeśli tak, w jaki sposób mogliśmy odziedziczyć cechę braku sierści po świniach?
Kiedy świnia ucieka z farmy i zaczyna żyć w lesie, nie staje się nagle zwierzęciem owłosionym. Jej potomkowie mogą, jeśli krzyżują się z owłosionymi dzikimi zwierzętami, ale nie w inny sposób. To prawda, że dzik eurazjatycki jest owłosiony (choć jego sierść nie jest tak gęsta jak np. u krowy czy owcy). Ale nie znamy historii świni domowej. Zwykle jest traktowana jako gatunek konspecyficzny z dzikiem euroazjatyckim, ale oba gatunki różnią się liczbą chromosomów (dla dzika domowego 2n=38, ale dla dzika 2n=36). Więc może się okazać, że nie są tym samym zwierzęciem i że stosunkowo bezwłose świnie podobne do świni domowej istniały w starożytności. Może się okazać, że oba gatunki były traktowane jako ten sam gatunek po prostu dlatego, że od dawna wiadomo, że mogą razem dawać płodne potomstwo. Ale to potomstwo może po prostu reprezentować hybrydy (to jeden z wielu przykładów zwierząt o różnej liczbie chromosomów, które razem dawały płodne potomstwo). Świnia domowa krzyżowała się również z wieloma innymi rodzajami świń, ale to nie oznacza, że są to te same zwierzęta. Na przykład, oprócz dzika, świnia domowa krzyżowała się z Babirusą, Babyrousa babyrussa ( zdjęcia ); świnią buszową, Potamochoerus larvatus ( zdjęcia ); świnią brodatą, Sus barbatus ( zdjęcia ); świnią brodawkowatą Visayan, Sus cebifrons ( zdjęcia ); dzikiem z Sulawesi, Sus celebensis ( zdjęcia ); i prawdopodobnie Sus oliveri i Sus philippensis . Dlaczego więc zakładać, że świnia domowa i dzik to „to samo” zwierzę? Stosunkowo nagie zwierzęta podobne do świni domowej mogły istnieć w starożytności. Tak naprawdę nie wiemy, jak wyglądały świnie tysiące lat temu, ale prehistoryczne malowidło w jaskini Altamira w Hiszpanii przedstawia świnię ( obrazy ), która wydaje mi się dość naga (poza tym, co wygląda jak broda i włosy na czubku głowy, szyi i ramionach).
Wideo: Brodata świnia
Niektóre z najłatwiej dostępnych dowodów, które można wykorzystać do oceny hipotezy hybrydowej, są widoczne w lustrze. W tej sekcji rozważymy pewne cechy zewnętrzne, które łączą ludzi ze świniami. Większość moich badań nad świniami koncentrowała się na zwykłej świni ( Sus scrofa ). Oczywiście, zwykła świnia to tak naprawdę ogólne określenie na wiele ras. „Obecnie na świecie istnieje około 87 ras świń domowych, większość z nich w Europie i Ameryce Północnej”, według Pond i Houpt, oraz „inne 225 lub więcej grup świń nieuznawanych za rasy, ale każda z nich ma unikalne cechy, wygląd lub lokalizację geograficzną”. 1 Jednak tutaj skupimy się na cechach, które są ogólnie charakterystyczne dla Sus scrofa .
A teraz przyjrzyjmy się bliżej pewnym ludzkim rozróżnieniom, które, jak się okazuje, są również cechami świń. Cechy, które odróżniają nas od szympansów i innych naczelnych, to jedyne, które zostaną omówione, ponieważ cechy, które ludzie dzielą z naczelnymi, nie mają znaczenia dla kwestii, czy ludzie są hybrydowego pochodzenia. Zgodnie z rozważaną hipotezą nie miałoby znaczenia, czy ludzie są bardziej podobni do szympansów pod wieloma względami niż do świń. Interesującym odkryciem jest to, że te cechy, które odróżniają ludzi od szympansów i innych naczelnych, można konsekwentnie wyjaśnić, odnosząc się do świni.
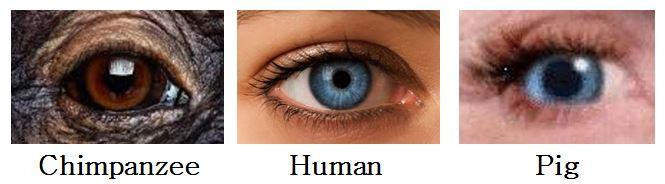
To fizyczne pokrewieństwo ludzi i świń jest łatwo zauważalne w pewnych cechach zewnętrznych. Fakt ten nie umknął Thomasowi Mannowi (1955, str. 271), który kiedyś napisał: „Świnia z małymi niebieskimi oczami, rzęsami i skórą ma więcej ludzkich cech niż jakikolwiek szympans — pomyśl, jak często nagie istoty ludzkie przypominają nam świnie”. Chociaż nie zgadzam się z twierdzeniem Manna, że świnie mają więcej wspólnych cech z ludźmi niż szympansy, uważam, że świnie i ludzie mają więcej niż wystarczająco wspólnych cech, aby sugerować związek. Na przykład, jasnopigmentowane oczy, w odcieniach niebieskiego, zielonego i jasnobrązowego, nigdy nie występują u szympansów lub orangutanów (Schultz 1947, str. 11). Najwyraźniej znany jest tylko jeden przypadek goryla z niebieskimi oczami (Dixson 1981, str. 31). Jasne oczy są również rzadkie u innych naczelnych ( tamże ). Dlaczego więc są one powszechne w niektórych populacjach ludzkich? Skąd wzięła się ta cecha? Jednym z możliwych wyjaśnień jest to, że została odziedziczona po niebieskookich świniach. Niebieski to powszechny kolor oczu u świń (podobnie jak zielony, żółty i jasnobrązowy). Ciemny pigment (melanina), tak powszechnie występujący w tęczówkach naczelnych innych niż człowiek, jest korzystny. Absorbuje światło ultrafioletowe . Aby chronić oczy przed tymi szkodliwymi promieniami, świnie polegają na swoich wąsko rozciętych, mocno rzęsionych powiekach. Ludzie osłaniają swoje oczy w podobny sposób, w przeciwieństwie do typowych małp o szeroko otwartych oczach i rzadkich rzęsach. [Czytelnik o nazwisku Chase Dumont napisał następujący komentarz, który jest interesujący w obecnym kontekście: „Wygląd zewnętrzny oka jest zupełnie inny niż u szympansa, bardziej przypomina oko świni — źrenica/tęczówka w oku szympansa pokrywa całe oko, podczas gdy w oku świni źrenica/tęczówka zajmują znacznie mniejszy obszar, odsłaniając większą część „białka” oka — podobnie jak u ludzi).”]
Nie jest też jasne, w jaki sposób wystający chrzęstny nos mógł pomóc wczesnym ludziom w ich „styl życia myśliwego na sawannie”. Jak zauważa Morris (1967, str. 67), „Ciekawe jest, że wystający, mięsisty nos naszego gatunku jest kolejną unikalną cechą, której anatomowie nie potrafią wyjaśnić”. Cecha ta nie jest charakterystyczna ani dla małp człekokształtnych, ani nawet dla innych wąsatych (Hill 1966, str. 29). Oczywiste jest, że świnie mają nos jeszcze bardziej wystający niż nasz. W ryju świni skrzydełka nosowe i przegroda są chrzęstne, tak jak u nas (Walker 1983, str. 1175). Natomiast nos szympansa „jest mały, płaski i nie ma bocznych chrząstek” (Sonntag 1923, str. 397). Chrzęstny nos jest najwyraźniej rzadką cechą u ssaków. Prymatolog Jeffrey Schwartz posuwa się nawet do stwierdzenia, że „to powiększona chrząstka skrzydełka nosowego sprawia, że ludzki nos jest tym, czym jest, i odróżnia ludzi od wszystkich innych zwierząt” (Schwartz 1987, s. 185). Chrzęstną strukturę ryja świni uważa się powszechnie za „adaptację” do kopania nosem (rycia). Rycie jest najwyraźniej wzorcem zachowania charakterystycznym dla świń. Inne zwierzęta kopią nogami.
Wystający nos jest prawdopodobnie najbardziej widoczną różnicą między twarzą człowieka a twarzą szympansa, ale dyskusje na temat ewolucji człowieka rzadko wspominają o nosie, być może dlatego, że jego brak użyteczności uniemożliwia wyjaśnienie w kategoriach adaptacji. Zamiast tego większość analiz zajmuje się bezmięsną czaszką, gdzie wystający nos człowieka jest nieco mniej oczywisty (ale mimo to widoczny). To osobliwe pominięcie, ponieważ bezużyteczne (nieadaptacyjne) cechy są powszechnie uważane za najlepsze wskaźniki pokrewieństwa. Jaka jest ewolucyjna użyteczność naszej unikalnej struktury nosa? Czy jest ona funkcjonalna? A może jest to pozostałość genetyczna starożytnego krzyża hybrydowego?
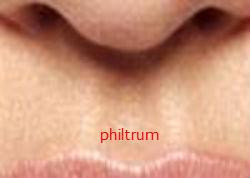
Kolejną cechą, którą należy wziąć pod uwagę, jest rynienka podnosowa, wgłębienie widoczne na środku górnej wargi człowieka. Małpy nie mają tej typowej ludzkiej cechy. 11 Wydaje się to bezużyteczną strukturą z punktu widzenia przetrwania. Dlaczego więc jest widoczna na całym świecie u Homo ? Zarówno u ludzi, jak i u świń, w początkowych stadiach rozwoju górna warga jest rozszczepiona, chociaż nie byłem w stanie znaleźć żadnych dowodów na taki rozszczep u zarodków żadnego naczelnego innego niż człowiek. W miarę postępu rozwoju rozszczep ten zwykle zamyka się u ludzi, ale utrzymuje się u świń. 12 Ludzka rynienka podnosowa jest widoczną pozostałością tej pierwotnej rozszczepionej wargi. U tych ludzi, u których ten rozszczep nigdy się nie zamyka, stan ten jest znany jako rozszczep wargi, powszechna wada wrodzona. Częste występowanie rozszczepu wargi u ludzi jest trudne do wyjaśnienia, jeśli założymy, że jesteśmy blisko spokrewnieni tylko z naczelnymi. Jeśli jednak założymy, że ludzie pochodzą ze skrzyżowania świni z szympansem, to odkrycie staje się o wiele bardziej zrozumiałe.
@Macroevo Szczerze mówiąc, hipoteza świni-szympansa jest po prostu genialna. Zastosowany zdrowy rozsądek sprawia, że zastanawiam się, jak nikt tego nie zauważył. Niesamowite
— alfredo romeo (@aromeo) 1 września 2013 r.
Podobne myślenie wyjaśnia krótkość górnej wargi człowieka (odległość między otworem gębowym a nozdrzami). Dlaczego nasza górna warga stała się krótsza i grubsza w toku ewolucji? Wszystkie małpy człekokształtne mają górne wargi znacznie dłuższe niż u ludzi, 13 ale górna warga świni jest tak krótka, że jest ledwie czymś więcej niż wyrostkiem ryja.14 Morris15 wiele uwagi poświęca faktowi , że ludzkie wargi są pokryte na swojej zewnętrznej powierzchni nagą (tj. całkowicie pozbawioną włosów) błoną śluzową :
Dalej sugeruje, że nasze osobliwe usta są produktem „doboru płciowego”. Ale można sobie wyobrazić inne wyjaśnienia: opisując skórę świń, Getty 16 stwierdza, że „nie ma prawdziwych nagich powierzchni poza krawędziami wargowymi”, które składają się z czerwonej błony śluzowej.
W odniesieniu do płatków uszu człowieka Morris zauważa, że „anatomowie często określają je jako bezsensowne wyrostki lub „bezużyteczne tłuszczowe narośla”. Niektórzy tłumaczą je jako „pozostałości” z czasów, gdy mieliśmy duże uszy. Ale jeśli przyjrzymy się innym gatunkom naczelnych, okaże się, że nie posiadają one mięsistych płatków uszu. Wydaje się, że są czymś nowym, dalekim od pozostałości”. 17 Być może jednak są czymś starym na nowej twarzy. Sisson opisuje dolną część ucha świni jako „mocno wypukłą od dołu, tworzącą wyniosłość nieco analogiczną do płatka ucha ludzkiego”. 18
Należy tu wspomnieć o dodatkowej cesze ludzkiego ucha, guzku Darwina (patrz ilustracja Darwina poniżej). W swojej książce Descent of Man Darwin komentuje tę cechę, która czasami znajduje się na krawędzi ludzkiego ucha, którą opisuje jako „mały tępy punkt, wystający z do wewnątrz złożonego brzegu lub helisy… Punkty te nie tylko wystają do wewnątrz, ale często trochę na zewnątrz, tak że są widoczne, gdy głowa jest oglądana bezpośrednio z przodu lub z tyłu. Mają zmienną wielkość i nieco inną pozycję,
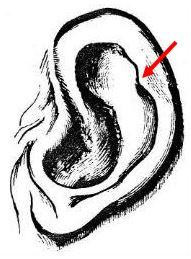
Guzek Darwina
(Darwin, 1871)
Prymatolog Adolph Schultz (1973) jednak stanowczo zaprzecza Darwinowi, mówiąc, że „wyraźnie spiczaste uszy, powszechnie nazywane 'uszami satyrowymi', są typowe dla małp tylko makaków i pawianów i nie występują u żadnych człekokształtnych [małp człekokształtnych], nawet we wczesnych stadiach rozwoju. Nie ma zatem żadnego uzasadnienia, aby interpretować okazjonalne 'guzki darwinowskie' na ludzkich uszach jako atawistyczny przejaw przodków o spiczastych uszach”. 20 Ale Schultz prawdopodobnie nie wziął pod uwagę spiczastych uszu świń.
Świnie mają wyraźne włoski brwiowe. Na brwiach płodu szympansa można dostrzec obszar jasnych guzków, układających się w wzór podobny do ludzkiego brwi. Dorosłe małpy nie mają jednak włosków brwiowych. 21 Na powiekach świnie mają bujne rzęsy, grubsze nawet niż u ludzi. U wielu świń te rzęsy , jak nazywają je anatomowie, są tak grube, że zwierzę wydaje się nosić sztuczne rzęsy. Jednak małpy człekokształtne prawie w ogóle nie mają rzęs, pomimo pozornej wartości przetrwania tej cechy. Ponadto pongidy mają wyraźne łuki brwiowe, podczas gdy świnie i większość ludzi ich nie mają. Jeśli zdecydujemy się wyjaśnić rozwój ludzkich rzęs i brwi w kategoriach doboru naturalnego, musimy się zastanowić, dlaczego małpy człekokształtne, które istniały co najmniej tak długo, jak jakikolwiek człowiekowaty, nie nabyły ich. Być może ich ciężkie łuki brwiowe wystarczająco chroniły ich oczy, ale jeśli tak jest, dlaczego łuki brwiowe nie wystarczyły również dla Homo ? Jaka pilna potrzeba skłoniła Homo do zastąpienia łuków kości kępkami włosów?
Charakterystyka skórna
W związku z hipotezą, że ludzkie pochodzenie można prześledzić do krzyżówki hybrydowej, ważne jest, aby zdać sobie sprawę, że w większości krzyżówek hybrydowych ssaków, męskie hybrydy są zazwyczaj bardziej bezpłodne niż żeńskie. Ten fakt oznacza, że hodowcy pracujący z hybrydami zazwyczaj łączą płodne samice z jednym z dwojga rodziców (czyli „krzyżują je wstecznie”). Z reguły nie tworzą nowych ras poprzez krzyżowanie hybryd z pierwszego krzyżowania między sobą.
Często, nawet po krzyżowaniu wstecznym, powstałe hybrydy są nadal płodne tylko w jednej płci. Tak więc zazwyczaj dochodzi do powtarzanego krzyżowania wstecznego. Jednak po wystarczającej liczbie krzyżowań wstecznych często uzyskuje się płodne hybrydy obu płci, a nowa rasa może być odtąd utrzymywana poprzez kojarzenia między samymi hybrydami. Powtarzane krzyżowanie wsteczne jest zazwyczaj bardziej konieczne w przypadkach, gdy rodzice uczestniczący w pierwotnym krzyżowaniu są bardziej odlegle spokrewnieni i genetycznie niekompatybilni. Tak więc można się również spodziewać, że w przypadku nowych typów organizmów powstających poprzez naturalną hybrydyzację, krzyżowanie wsteczne będzie zwykłą drogą do płodności i stabilności reprodukcyjnej. I to samo miałoby miejsce w konkretnym przypadku ludzi powstających poprzez hybrydyzację.
Jednakże hybrydy krzyżowane wstecznie są trudne do zidentyfikowania za pomocą technik molekularnych i stają się trudniejsze z każdym kolejnym krzyżowaniem wstecznym ( Vähä i Primmer 2006 , Engler i in. 2015 ). Vähä i Primmer twierdzą, że aby wiarygodnie zidentyfikować nawet pierwsze krzyżowania wsteczne za pomocą konwencjonalnych technik, należy zbadać co najmniej 48 loci (a przy powtarzanym krzyżowaniu wstecznym liczba ta znacznie wzrasta). Aby lepiej zrozumieć ten problem, rozważmy hipotetyczny wpływ powtarzanego krzyżowania wstecznego na genom ludzki. Czytelnik może nie być zaznajomiony ze zjawiskiem konwersji genów , ale jego wpływ na hybrydy podczas krzyżowania wstecznego polega na szybkiej homogenizacji sekwencji genów. Aby zrozumieć, dlaczego tak jest, rozważmy wpływ krzyżowania wstecznego na hybrydowe DNA.
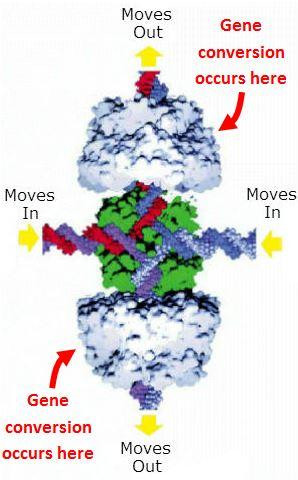 Złącze Hollidaya. Podczas rekombinacji mejotycznej podwójne nici rodzicielskiego DNA, pokazane tutaj w ruchu do złącza, rozdzielają się na dwie pojedyncze nici. Każda pojedyncza nić od jednego rodzica łączy się następnie z pojedynczą nicią od drugiego rodzica. Powstałe złożone podwójne nici wychodzą ze złącza, przechodząc w tym procesie konwersję genów.
Złącze Hollidaya. Podczas rekombinacji mejotycznej podwójne nici rodzicielskiego DNA, pokazane tutaj w ruchu do złącza, rozdzielają się na dwie pojedyncze nici. Każda pojedyncza nić od jednego rodzica łączy się następnie z pojedynczą nicią od drugiego rodzica. Powstałe złożone podwójne nici wychodzą ze złącza, przechodząc w tym procesie konwersję genów.Na powyższym rysunku zauważ, że w każdym z rodzicielskich (nadchodzących) podwójnych nici każdy nukleotyd w jednej nici jest prawidłowo sparowany z komplementarnym nukleotydem w drugiej nici, A jest zawsze sparowany z T, a C jest zawsze sparowany z G. Tak więc sparowana podwójna nić DNA od jednego rodzica może wyglądać następująco:
AG T TCCGACGCG
TC A AGGCTGGC
podczas gdy pasmo od drugiego rodzica może wyglądać tak:
AG C TCCGACGCG
TC G AGGCTGGC
W każdym z tych dwóch podwójnych łańcuchów wszystkie nukleotydy są sparowane z komplementarnym nukleotydem (A zawsze z T, a C zawsze z G). Ale gdy porównamy jeden podwójny łańcuch z drugim, widać wyraźnie, że różnią się one trzecią pozycją zasad nukleotydowych. W pierwszym podwójnym łańcuchu para zasad nukleotydowych to TA, podczas gdy w drugim jest to CG.
Tak więc, gdy te nici połączą się ze swoimi nowymi niciami partnerskimi po przejściu przez złącze Hollidaya w mejozie, dwa powstałe podwójne nici będą:
AG T TCCGACGCG
TC G AGGCTGGC
I
AG C TCCGACGCG
TC A AGGCTGGC
Tak więc T jest sparowane z G w pierwszym wychodzącym paśmie, a C jest sparowane z A w drugim wychodzącym paśmie. Konwersja genów przekształca każdą taką niedopasowaną parę w dopasowaną parę poprzez zastąpienie jednego z dwóch nukleotydów uzupełnieniem drugiego nukleotydu (który pozostaje niezmieniony). Wyniki eksperymentów sugerują, że mechanizm wybiera losowo, który z dwóch nukleotydów zastąpić, tak aby nukleotydy pochodzące od każdego z rodziców przeżyły w równym tempie.
Tak więc w dowolnym przypadku krzyżowania wstecznego załóżmy, że genomy pierwotnych rodziców A i B, którzy wyprodukowali hybrydy pierwszego krzyża, różniły się jedną pozycją nukleotydu na pięć (20%). Wtedy DNA w gametach hybryd różniłoby się od A jedną pozycją na dziesięć, a od B jedną pozycją na dziesięć. Oznacza to, że gamety pierwszego krzyża znajdowałyby się dokładnie pośrodku między A i B pod względem sekwencji genów.
Jednakże po jednym krzyżowaniu wstecznym z A i wynikającej z tego konwersji genu podczas mejozy w hybrydach krzyżowania wstecznego, gamety wytworzone przez osobniki krzyżowania wstecznego różniłyby się od A tylko w jednej pozycji na dwadzieścia (5%). A gamety wytworzone przez drugie krzyżowanie wsteczne różniłyby się tylko w jednej pozycji na czterdzieści (97,5% podobieństwa). Jest zatem jasne, że szybko staje się dość trudno odróżnić, na podstawie danych sekwencji nukleotydów, hybrydy krzyżowania wstecznego od czystego rodzica A, do którego doszło krzyżowanie wsteczne. Szympansy i ludzie są w około 98% podobni pod względem sekwencji nukleotydów.
Konkretne podstawy genetyczne licznych cech, które odróżniają ludzi od szympansów, wyjaśniono w kontekście niniejszej teorii w osobnej sekcji zatytułowanej Dlaczego ludzie różnią się od szympansów?
To, że ludzie nie mają okrywy włosowej, jaką mają naczelne inne niż człowiek, jest powszechnie akceptowanym faktem. „To właśnie ten pojedynczy czynnik stanowi główną różnicę między skórą ludzką a skórą innych ssaków” (Montagna 22 ). Niektórzy autorzy twierdzą, że okrywa włosowa szympansa jest „rzadka”. Ale jeśli „rzadka” opisuje okrywę włosową szympansa, to „naga” dokładnie opisuje skórę ludzi. Każdy człowiek, który choćby zbliżyłby się do owłosienia innych naczelnych, byłby uważany za nienormalnego. Świnie są jednak innym przypadkiem. Wiele ras świń domowych ma skórę tak samo nagą, jak skóra ludzka. Jak zauważają Cena i in . (101.9,521), „Gęstość owłosienia [w sierści zwierząt] waha się od rzadkiej resztkowej okrywy u człowieka i świni z 10-100 włosami na cm² do [gęstej] sierści gatunków takich jak lis i królik z około 4000 włosami na cm²”. Według Haltenortha, u dzikiego gatunku Sus scrofa gęstość owłosienia waha się od „rzadkiego do gęstego”, w zależności od okazu lub odmiany. 23 Na przykład, owłosienie współczesnej dzikiej odmiany Sus scrofa występującej w Sudanie ( S. s. senaarensis ) jest dość rzadkie. 24
Inne naczelne nie mają długiej grzywy włosów na czubku głowy niestrzyżonego człowieka, ani nie mają brody. Haltenorth zauważa, że u niektórych odmian Sus scrofa grzywy znajdują się na szyi i plecach, brody na policzkach, a kosmyki włosów na czole i na czubku głowy. Mówi również, że ostatnią z tych trzech cech można znaleźć u świń tylko u Sus . 25 Prehistoryczne malowidło świni znalezione w jaskini Altamira w północnej Hiszpanii przedstawia zwierzę z brodą i gęstymi włosami na czubku głowy ( zdjęcia ). Sus barbatus , istniejąca świnia pochodząca z Azji Południowo-Wschodniej (która tworzy płodne mieszańce obu płci w krzyżówkach z S. scrofa ), ma mało włosów na ciele, ale ma bardzo gęstą i krzaczastą brodę. 26
Panniculus adiposus. W artykule na temat ewolucji ludzkiej skóry, znany anatom porównawczy skóry William Montagna (1985, str. 14) zauważa, że „wraz z utratą futrzastej pokrywy, ludzka skóra nabyła podskórną warstwę tłuszczu ( panniculus adiposus ), która jest znacznie grubsza niż u innych naczelnych, czy ssaków. Nie oznacza to, że tylko człowiek ma tłustą skórę, ale gruba warstwa tłuszczu jest tak samo charakterystyczną cechą ludzkiej skóry, jak skóry świńskiej”. Podobnie Dyce i in . (1987, str. 742) zauważają, że istnieje „dobrze rozwinięty depozyt tłuszczu obecny niemal wszędzie w tkance podskórnej”. Prymatolog FW Jones (1929, str. 309) również zauważył tę warstwę tłuszczu:
Panniculus carnosus. „Inną szczególną cechą ludzkiej skóry jest ogólny brak lub utrata warstwy mięśni szkieletowych ( panniculus carnosus ), występującej w skórze większości innych ssaków. Pozostałości panniculus carnosus w ludzkiej skórze znajdują się z przodu szyi w przypominającym fartuch, cienkim mięśniu platysma… Wszystkie inne naczelne, nawet małpy człekokształtne, mają panniculus carnosus na większości ciała” (Montagna (1985, s. 14-15; patrz również Sobotta 1983, ryc. 459, 460). Podobnie jak u ludzi, muskulatura skórna świń jest dobrze rozwinięta na szyi (mięsień platysma) i twarzy, ale skąpa lub nieistniejąca w innych miejscach (Nickel i in. 1986, s. 230; Sack 1982).
U zwierząt mających podskórną tkankę tłuszczową , skóra otrzymuje zaopatrzenie w krew z bezpośrednich tętnic skórnych (duże naczynia powierzchniowe biegnące równolegle do powierzchni skóry w pochewce mięśniowej skóry). Ale gdy podskórna tkanka tłuszczowa nie występuje, tętnice zasilające skórę wznoszą się jak małe drzewa z głębi ciała. Tętnice tego ostatniego typu nazywane są mięśniowo-skórnymi . Te dwie formy krążenia skórnego przedstawiono na poniższej ilustracji. Zarówno skóra świni, jak i skóra człowieka są zaopatrywane przez tętnice mięśniowo-skórne (Montagna i Parakkal 1974, s. 143-144). Jak zauważyli Daniel i Williams (1973, str. 20-21) w artykule na temat transferu płata skórnego, „Większość zwierząt doświadczalnych nie ma unaczynienia skóry podobnego do ludzkiego. Wykazano anatomicznie i chirurgicznie, że unaczynienie skóry świni jest bardziej porównywalne niż większości u człowieka… Podobnie jak u człowieka, skóra świni jest unaczyniona przez wszechobecne tętnice mięśniowo-skórne i przez kilka bezpośrednich tętnic skórnych”. Obserwację tę potwierdzili inni autorzy: „Z wyjątkiem świń, których unaczynienie skóry przypomina ludzkie, ssaki o luźnej skórze są unaczynione przez bezpośrednie tętnice skórne” (Montagna i Parakkal 1974, str. 144). Dlatego pod tym względem skóra ludzka jest bardziej podobna do skóry świńskiej niż do skóry naczelnych: „Właściwie unaczynienie skóry większości naczelnych jest zasadniczo podobne do unaczynienia skóry innych zwierząt futerkowych” (Montagna 1985, s. 16). W szczególności Baccaredda-Boy (1964, s. 211), a także Moretti i Farris (1963, s. 162, 170-171) odkryli, że skóra szympansów różni się od skóry ludzi tym, że ma liczne duże, powierzchowne naczynia (tj. bezpośrednie tętnice skórne).
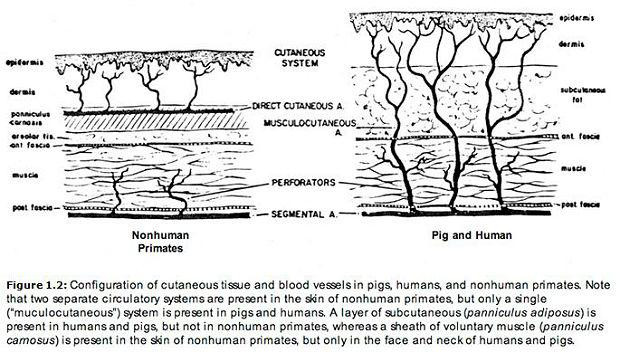
Skóra człowieka wyróżnia się także od skóry innych naczelnych — a także większości innych ssaków — ilością krwi, która może przez nią krążyć (Montagna 1985). Pewna ilość krwi jest potrzebna tylko do odżywienia skóry. Jest to ilość, którą otrzymuje ona u większości zwierząt. U ludzi jednak maksymalny przepływ krwi może być ponad sto razy większy od tego minimalnego (Burton 1961; Montagna i Parakkal 1974, s. 151; Rowell 1977, s. 155). Odżywiane przez wrażliwe na temperaturę tętnice mięśniowo-skórne, gęsto rozmieszczone naczynia włosowate skóry u ludzi odgrywają istotną rolę termoregulacyjną (Montagna 1985, s. 16). Kiedy ciało zaczyna się przegrzewać, duże ilości ciepłej krwi mogą zostać szybko schłodzone w tych naczyniach włosowatych poprzez parowanie potu. Jednym z mierników gęstości naczyń skórnych jest odstęp między pętlami naczyń włosowatych. U ludzi typowa odległość między naczyniami włosowatymi waha się od 50 do 100 mikronów (Ryan 1973, str. 619). W skórze boków świń ta wartość jest zredukowana do zaledwie około 20 mikronów, odstęp ten jest tak mały, że aż niewiarygodny. Kiedy białe świnie są wystawione na działanie wysokich temperatur, ich skóra czerwienieje od krwi (nawet przy braku światła słonecznego), tak jak dzieje się to u ludzi o jasnej karnacji w podobnych warunkach (Roller i Goldman 1968, str. 13).
 |
Pchła ludzka, Pulex irritans |
Pchły. Być może ta różnica między naszym układem naczyniowym skóry a układem naczyń krwionośnych naszych naczelnych, bogactwo uczty pod naszą skórą, wyjaśnia inną ludzką odmienność: „Ironicznie”, pisze Nicole Duplaix (1988, str. 694), „człowiek jest wyjątkowy wśród naczelnych, ponieważ ma pchły”. Ponad 2400 odrębnych typów pcheł zostało potraktowanych jako gatunki lub podgatunki ( tamże , str. 689). Pasożyty są zazwyczaj dość specyficzne w wyborze żywiciela. Mniej niż dwadzieścia z tych 2400 typów z łatwością ugryzie ludzi ( tamże , str. 685). Najważniejszym z tych, które żywią się Homo sapiens, jest pchła ludzka, Pulex irritans , ale nie jesteśmy jedynymi odpowiednimi żywicielami dla tego gatunku. Według Bennetta (1975, s. 775) „ Pulex irritans , pchła ludzka, rozmnaża się swobodnie w ściółce chlewni i może stać się poważnym szkodnikiem świń”.
Poduszeczka tłuszczowa zastępuje włosy jako warstwę izolacyjną u ludzi i świń. Według Becketta (63.8,2),
Na powyższym rysunku zauważ, że tętnice mięśniowo-skórne przechodzą przez tłuszcz skórny. Ta perforowana warstwa tłuszczu stanowi mechanizm izolacyjny, który może szybko reagować na temperaturę otoczenia, cechę, której brakuje włosom. Rozszerzenie tętnic mięśniowo-skórnych w odpowiedzi na ciepło zwiększa przepływ krwi do skóry. Ten wzrost krążenia może podnieść temperaturę powierzchni skóry do poziomu prawie tak wysokiego, jak temperatura w ciele, zwiększając w ten sposób szybkość, z jaką ciepło jest tracone do otoczenia. b W chłodnym otoczeniu zwężenie tych tętnic obniża temperaturę skóry, a w konsekwencji szybkość, z jaką ciepło ciała jest tracone do atmosfery, ponieważ warstwa tłuszczu może wówczas służyć jako koc izolacyjny.
Posiadanie tkanki tłuszczowej pozwala na dostosowywanie się do zmian temperatury otoczenia w każdej chwili — jest to wyraźna zaleta w strefach umiarkowanych, w których osiedliła się znaczna część rasy ludzkiej, ponieważ regiony te są znacznie bardziej podatne na nagłe, ekstremalne zmiany temperatury niż te w pobliżu równika. Naczelne inne niż człowiek i inne zwierzęta futerkowe nie mają możliwości dostosowywania temperatury skóry. Ponieważ ich skóra nie jest izolowana od reszty ciała warstwą tłuszczu, jej temperatura musi utrzymywać się w pobliżu temperatury ciała pod nią.
Skóra świni jest oddzielona od wnętrza ciała grubą warstwą tłuszczu i może się wychładzać do ekstremalnego stopnia. Tłuszcz, a nie włosy, jest główną barierą izolacyjną. 47 Świnie alaskańskie mogą wytrzymać temperatury poniżej zera, schładzając swoją skórę do zaledwie 9℃ (przy temperaturze otoczenia -10℃) bez uszkodzenia tkanek. 48 Zaaklimatyzowani ludzie również mogą obniżyć temperaturę skóry do około 10℃ bez obrażeń. 49 Ten sposób izolacji jest zupełnie inny niż u naczelnych, bardziej przypomina ten obserwowany u niektórych ssaków wodnych (np. fok, morsów). Z wyjątkiem świni, wydaje się, że żadne inne zwierzę lądowe nie ma takiej formy izolacji.
Więcej niż naga małpa, Homo jest nagą małpą o zmiennej izolacji. W gorącym środowisku ludzie (i świnie) mogą zwiększyć krążenie ciepłej krwi do skóry i podnieść temperaturę prawie do poziomu temperatury rdzenia ciała, maksymalizując w ten sposób utratę ciepła do otaczającego powietrza. Jeśli pogoda się ochłodzi, mogą ograniczyć krążenie skórne, chłodząc skórę do takiego stopnia, że utrata ciepła jest znacznie zmniejszona. Ta zdolność jest szczególnie widoczna u osób otyłych 50 lub zaaklimatyzowanych. 51 Chociaż postęp kulturowy, wynalezienie odzieży, umożliwił Homo zamieszkiwanie chłodnych regionów, które wcześniej były niedostępne dla naczelnych, postęp biologiczny w postaci nowego systemu izolacji zwiększył ludzką zdolność do wytrzymywania nagłych zmian temperatury występujących w tych regionach.
Oprócz tego, że jest dobrym izolatorem, ludzka skóra jest zaskakująco gruba. „Naskórek na naszej ogólnej powierzchni ciała [„ skóra owłosiona ” patrz uwaga po prawej] jest znacznie grubszy niż u innych naczelnych: warstwa rogowa [warstwa rogowa] może być odklejona w stanie nienaruszonym jako przezroczysta, ale wytrzymała błona, która może być używana do celów eksperymentalnych… Naskórek w skórze owłosionej u naczelnych innych niż człowiek, podobnie jak u każdego innego ssaka futrzanego, jest stosunkowo cienki, z stosunkowo cienką warstwą rogową” (Montagna 52 ). Świnie mają jednak gruby naskórek i warstwę rogową, grubsze nawet niż u ludzi. 53
Elastyczność naszej skóry jest również niezwykła. „Podczas gdy skóra wielkich małp i niektórych małpich naczelnych ma zmienną ilość włókien elastycznych, u żadnych zwierząt, niezależnie od płci, wieku lub miejsca występowania, nie znaleźliśmy obfitości tkanki elastycznej charakterystycznej dla skóry ludzkiej” (Montagna 54 ). Odkrycie to pochodzi od tego samego autora, który we wcześniejszym artykule porównującym skórę ludzką ze skórą świń, zauważył, że „jednym z najbardziej uderzających podobieństw między tymi dwiema skórami [świńską i ludzką] jest duża zawartość tkanki elastycznej w skórze właściwej”. 55
Zauważa również, że „powierzchnia obu skór [ludzkiej i świńskiej] jest wyżłobiona przecinającymi się liniami, które tworzą charakterystyczne wzory geometryczne”. 56 W osobnym artykule na temat ewolucji skóry ludzkiej podaje nieco więcej szczegółów:
Obecność tych linii u świń i ludzi nie jest łatwa do wyjaśnienia w kategoriach doboru naturalnego, ponieważ nie mają one żadnej znanej funkcji. 58
Na spodniej stronie naszej „włochatej skóry” (ogólna powierzchnia ciała), gdzie naskórek styka się ze skórą właściwą, znajduje się inny wzór, który nie odpowiada w swojej konfiguracji wzorowi zewnętrznemu opisanemu w poprzednim akapicie. Podobny, choć grubszy wzór jest również charakterystyczny dla połączenia naskórkowo-skórnego u świń. Montagna zauważa jednak, że „w preparatach skóry rozdzielonej, gdzie naskórek jest starannie usuwany ze skóry właściwej, naskórek zwierząt o dużej sierści jest płaski. 59 Nawet u małp i małp człekokształtnych bruzdy naskórkowe znajdują się tylko wokół przyłączenia przewodów gruczołowych i kanałów włosowych”. Możemy wyjaśnić drobniejszy wzór u ludzi niż u świń faktem, że drobna siatka jest pośrednia między grubym wzorem skóry świńskiej a gładką spodnią powierzchnią skóry naczelnych innych niż człowiek.
Tak więc u świni mamy zwierzę o rzadkiej sierści, z tłustą, rozciągliwą skórą, do której doprowadzają tętnice mięśniowo-skórne. Powierzchnia owłosionej skóry jest oznaczona wrodzonymi liniami podobnymi do tych, które można zaobserwować u ludzi, a wzór połączenia naskórkowo-skórnego jest również bardzo podobny w regionach owłosionej skóry. Zgodnie z hipotezą, którą rozważamy, nie ma większego znaczenia, że skóra świni różni się od skóry ludzkiej w innych kwestiach. Istotnym punktem jest to, że w przypadkach, w których nasza skóra jest specyficzna dla naczelnych, wyjaśnienie każdej takiej anomalii można znaleźć w skórze świń.
Łowca Sawanny
Dojrzała świnia ma około 500 000 dużych gruczołów potowych rozmieszczonych na całym ciele (503,3497; 506,5316). Niemniej jednak w literaturze często twierdzi się, że świnie się nie pocą. Założenie to można prześledzić do badań Ingrama i Mounta, którzy badali wskaźniki pocenia u niedojrzałych zwierząt, zwykle u prosiąt prowadzących siedzący tryb życia (247,03; 247,1; 389,7; 390,1; 390,2; 390,3; 390,5). Badania oceniające pocenie się świń koncentrowały się na młodych świniach, ponieważ są one bardziej interesujące pod względem komercyjnym. Niedojrzałe zwierzęta nie są bardziej odpowiednie do określania właściwości parowania knura lub lochy niż małe dziecko do ujawniania cech dorosłego człowieka — dzieci pocą się znacznie mniej niż dorośli (584,4577). Małe zwierzęta mają tendencję do hipotermii (ponieważ ich powierzchnia jest duża w stosunku do ich rozmiaru), nie do hipertermii i mają niewielką tendencję do pocenia się (390.8,182). Pocenie się u świń jest często pomijane, ponieważ zwierzęta te najwyraźniej pocą się bardziej wydajnie niż ludzie. Ich gruczoły potowe wydają się być lepiej dostrojone do potrzeb termoregulacyjnych (nie produkują więcej potu niż jest to konieczne do schłodzenia krwi skórnej przez parowanie). Bardzo mało potu jest tracone do odpływu, ponieważ pot rzadko gromadzi się na skórze. Jednak obserwowane wskaźniki pocenia się u dojrzałych świń są w przybliżeniu porównywalne do wskaźników u ludzi. Beckett (63.4) stwierdził, że 350-funtowa locha w stanie spoczynku traciła około 95 g/m² potu na godzinę przy temperaturze suchego termometru 98℉ i temperaturze mokrego termometru 81℉. W znacznie wyższej temperaturze (122℉ suchy termometr i 79℉ mokry termometr) Myhre i Robinson odkryli, że 70-kilogramowi mężczyźni w stanie spoczynku tracili wilgoć (pot + oddychanie) w tempie 250 g/m² na godzinę (398,7, Tabela 3). Nawet u mniejszych świń (loszek o wadze 198 funtów) utrata wilgoci ze skóry jest ważna (387,8, Tabela 1), stanowiąc od jednej trzeciej do dwóch trzecich całkowitej utraty wilgoci (płuca + skóra). Pojawia się również twierdzenie, że świnie potrzebują wanienki, gdy żyją w gorącym klimacie (ponieważ rzekomo się nie pocą). Jednak Heitman i Hughes wystawili świnie bez dostępu do wanienki na wysokie temperatury (100℉ względna wilgotność 35%) przez tydzień bez żadnych ofiar śmiertelnych — warunki, w których jedyną drogą do rozproszenia ciepła jest chłodzenie parowe (232,5,176).
Świnie pocą się, gdy jest im gorąco. „Apokrynowe [tj. potowe] gruczoły u koni i świń wydzielają obficie podczas gwałtownych ćwiczeń i stresu” (Montagna 60 ). To pocenie pełni funkcję termoregulacyjną u świń, tak samo jak u ludzi. 61 Owłosione gruczoły potowe naczelnych nie reagują jednak na stymulację cieplną. Niezdolność naczelnych do pocenia się wprawiła Montagnę w zakłopotanie: „Można by przypuszczać”, pisze,
Natomiast nawet mała dawka acetylocholiny lub adrenaliny wywołuje u świń pocenie się.63 Nawet niedojrzałe świnie badane przez Ingrama (247.1,95) reagowały na adrenalinę.
Powszechnie przyjmuje się pogląd, że nagość w jakiś sposób zwiększyła parowanie potu u ludzi. Podobno nasza rzadka sierść pozwalała naszym przodkom chłodzić skórę szybciej niż owłosione zwierzęta w gorącym, suchym środowisku lub w jakiś sposób poprawiała ich zdolność do rozpraszania ciepła metabolicznego podczas biegania po sawannie w pogoni za zdobyczą. Russell Newman wskazuje jednak, że nasz brak odbijających włosów faktycznie zwiększa obciążenie ciepłem słonecznym i potrzebę pocenia się. 164 Aby uzasadnić to twierdzenie, cytuje badanie Bermana, które wykazało, że bydło wystawione na słońce poci się bardziej po usunięciu sierści. 165 Podobnie, dyszenie nasila się u strzyżonych owiec. 166
Ubrania, które zastępują włosy jako barierę przed promieniowaniem u ludzi, mają w zasadzie taki sam wpływ na ludzkie pocenie się. Ludzie poddani obciążeniom słonecznym pocą się bardziej, gdy są nadzy, niż gdy noszą lekkie ubrania w identycznych okolicznościach. W badaniu wpływu ubrań na pot Adolph 167 doszedł do wniosku, że „nagi mężczyzna może zaoszczędzić tyle samo wody w organizmie, zakładając koszulę i spodnie, co ubrany mężczyzna, znajdując dobry cień”. Ponadto owłosienie ciała nie zmniejsza utraty ciepła konwekcyjnego „i nie ma nic wspólnego z promieniowaniem długofalowego ciepła podczerwonego do chłodniejszych obiektów”, mówi Newman. 168 Dlatego twierdzi, że naga skóra,
Jeśli zwiększone obciążenia cieplne promieniowania sprawiły, że wczesni ludzie bardziej polegali na pocie w celu chłodzenia, dlaczego utrata włosów, która zwiększa te obciążenia, postąpiła w takim stopniu, jak u Homo ? Zakładając, że ludzie najpierw ewoluowali na suchej, nasłonecznionej sawannie, trudno jest, pod względem wydajności przetrwania, wyjaśnić zmniejszenie gęstości włosów, które skutkowałoby zwiększonym tempem spożycia wody. Newman wskazuje, że nie ma dowodów na to, że włosy zakłócają parowanie potu. Właściwie sam przeprowadziłem prymitywny eksperyment, którego wyniki wskazują, że włosy faktycznie przyspieszają parowanie potu. Odkrycie to jest zaskakujące w świetle częstych twierdzeń teoretyków ewolucji, że jest inaczej. Ale po odrobinie zastanowienia można zdać sobie sprawę, że sierść nie jest barierą parową. Zdolność futra do „oddychania” zawsze odróżniała je od mniej pożądanych izolatorów, które spowalniają utratę ciepła, ale nie „odprowadzają” wilgoci ze skóry. Dlaczego włosy nie tylko powinny umożliwiać, ale nawet zwiększać tempo parowania? Istnieją co najmniej dwa powody. Po pierwsze, mokre włosy mają bardziej nieregularną powierzchnię w stosunku do otaczającej atmosfery niż skóra bez włosów, co zwiększa powierzchnię dostępną do parowania. Po drugie, włosy umożliwiają równomierne rozprowadzanie potu poprzez działanie kapilarne, zapobiegając tworzeniu się pojedynczych kropelek widocznych na nagiej skórze. Gdy takie kropelki się tworzą, skóra leżąca między nimi nie służy jako powierzchnia parowania, a szybkość parowania jest zmniejszona.
W miarę jak ilość potu na skórze wzrasta, poszczególne krople łączą się, tworząc ciągły arkusz wody. Ale gdy na nagiej skórze znajduje się duża ilość potu, pojawia się inny rodzaj nieefektywności — odpływ. Więcej potu spływa z bezwłosej skóry bez odparowywania. Sierść owłosionego zwierzęcia działa jak gąbka, zatrzymując pot w miejscu, aż do momentu, gdy będzie mógł odparować. Pot spływający z ciała nie ma efektu chłodzącego, ponieważ odpływ nie pochłania ciepła. Natomiast odparowujący pot pochłania dużą ilość ciepła. 1a Jednak badania Adolpha wskazują, że około jednej czwartej ludzkiego potu może zostać utracona w wyniku odpływu, nawet w warunkach niemal optymalnych parowania. 1b Odbijająca sierść ma zatem trzy zalety: (1) mniejsze obciążenie ciepłem słonecznym; (2) zwiększone tempo parowania; (3) mniej potu marnowanego na odpływ. Trudno zatem zrozumieć, w jaki sposób nagą skórę można interpretować jako „adaptację” korzystną dla myśliwego na sawannie.
Oczywiście hipoteza „łowcy sawanny” jest tylko jedną z wielu teorii. Wypadanie włosów u Homo było przedmiotem wielu spekulacji (przegląd takich teorii można znaleźć w 165.1). Oprócz tych, którzy twierdzą, że straciliśmy włosy na sawannie 170 i/lub dlatego, że byliśmy myśliwymi 171 , są inni, którzy sugerują, że mogliśmy je stracić w lesie 172 lub nawet w morzu. 173 Niektórzy autorzy sugerują, że nagość czyniła nas seksualnie kuszącymi 174 lub że brak owłosienia stał się korzystny termicznie, gdy zaczęliśmy nosić ubrania. 175
Nawet gdybyśmy chcieli założyć, że ludzie kiedyś mieli sierść (nie ma absolutnie żadnych dowodów na to, że tak było), te teorie nie wyjaśniałyby zalet rzadkiej sierści. Hipoteza łowiecka jest nie do utrzymania, ponieważ wszystkie drapieżniki lądowe inne niż człowiek mają gęstą sierść. Podobne zastrzeżenie można podnieść w odniesieniu do scenariusza seksualnego pokusy. Dlaczego nie wszystkie ssaki straciły sierść, skoro nagość jest kusząca? Propozycja wodna jest również wątpliwa, większość małych (wielkości człowieka lub mniejszych) ssaków wodnych lub półwodnych ma sierść. 176
Hogu I
Amo, małpa przerażona
Hic haec, świnia czkawka Świnia,
łaciński kochanek
Hogu II
Amo amas amat
Małpa przerażona agape Świnia,
łaciński kochanek
Hogu III
Małpa przeciwko świni
Agape, agape, agape
Świnia, grecki kochanek
Hogu IV
Małpa, agape i świnia
Raz za razem i jeszcze raz, i jeszcze raz
Potem człowiek
Hogu V Małpa,
agape
Świnia i szturchnięcie
Vo-lar-e, kto-ho-ho-świnia
Hogu VI
Bez świni nie ma zysku
Małpa, agape, świnia znowu
Świnia w małpie nieludzki
Hogu VII
Świnia w małpiej przepaści
Fałszywy orgazm
Aktor szynki
Hogu VIII
Bezczelne chińskie szympansy
Jak świnia w składzie porcelany
Porcelana
Wyniki mojego eksperymentu z parowaniem utrudniają mi zaakceptowanie opinii Mounta, że naga skóra odparowuje pot szybciej niż skóra owłosiona. 177 Z tego samego powodu wątpię w sugestię Wheelera, że nabycie wyprostowanej postawy przez hominidy „było prawdopodobnie podstawową preadaptacją, która umożliwiła im zrzucenie owłosienia ciała i rozwinięcie rozległych powierzchni parowania”. 178 Również wątpliwa jest „hipoteza westiariusza” Kushlana, ponieważ zakłada ona, że wynalezienie odzieży pozostawiło Homo swobodę w utracie owłosienia ciała i uzyskaniu w ten sposób „najbardziej wydajnego systemu chłodzenia spośród wszystkich ssaków”. 179 Jak widzieliśmy, naga skóra nie zapewnia żadnej szczególnej przewagi parowania.
Ponieważ nagość jest przeszkodą na sawannie, Newman dochodzi do wniosku, że mało prawdopodobne jest, aby przodkowie człowieka tracili włosy po opuszczeniu lasu: „Gdybyśmy musieli wybrać okresy, w których postępująca dendacja nie stanowiła wyraźnej przeszkody środowiskowej, wybór byłby następujący:
Należy jednak zauważyć, że Newman nie wyjaśnia, dlaczego nasi przodkowie tracili włosy w tym samym środowisku (lesie), w którym małpy nie traciły włosów. Jeśli ludzie powstali poprzez krzyżowanie świń i szympansów, ich geneza prawie na pewno miała miejsce w lesie. Szympansy żyją w lasach. Na podstawie wysokiego wskaźnika zużycia wody Yang wywnioskował, że świnia również jest funkcjonalnie zwierzęciem leśnym. 182 Ludzie potrzebują więcej wody niż prawie każde inne zwierzę. 183
Rzeczywiście, wydaje się nieprawdopodobne, aby człowiekowaty spędzał więcej czasu niż to konieczne poza lasem. Chociaż sawanny Afryki roiły się od zwierzyny, roiło się tam również od dzikich drapieżników. Kiedy człowiek jest ścigany przez lwa, pierwszym impulsem jest znalezienie drzewa. Rozważ obraz malowany przez obecną teorię ewolucji: szlachetny myśliwy z sawanny, nagi wobec bezczelnego słońca, śmiało wyprostowany na jałowej i bezdrzewnej równinie, w niestrudzonym pościgu za ostrożną i niebezpieczną zdobyczą, obojętny na atak drapieżnych drapieżników. Z pewnością ten opis ma dramatyczny urok. To jak opowieść o Tarzanie. Ale czy jest prawdopodobny?
Ta przodkowa zmiana jest duża.
Nie tylko małpy mogą rościć sobie prawo do tego przydziału.
Właśnie odkryłem,
skąd wziąłem swój wielki ryj.
Okazuje się, że pradziadek był świnią!
Powyższy limeryk jest czytany przez Carla Kasella w programie National Public Radio Wait Wait... Don't Tell Me! Wyjaśnienie Petera Sagala: „Cóż, genetyk wykształcony na University of Georgia wysunął teorię, że rasa ludzka narodziła się, gdy szympansica zajęła się świnią. A to dlatego, że ludzie i świnie mają zaskakująco wiele wspólnych cech: pozbawioną sierści skórę, bardzo rzadką u naczelnych, jasne oczy i miłość do bekonu”.
Świetny artykuł: Genetyk z Georgii kwestionuje ewolucję, łączy ludzi ze świniami http://t.co/JIFFlR2JRs za pośrednictwem @sharethis — luca turin (@lucaturin) 29 lipca 2013
|
CYTATY:
1. (443,6,14)
2. (331,35,271)
3. (495.1,11)
4. (140.1,31)
5. (140.1,31)
6. (385,5,67)
7. (235.4,29)
8. (588,4,1175)
9. (533,6397)
10. (505.1,185)
11. (533,8,103)
12. (530.9,Ryc. 502)
13. (285,5,86)
14. (525,3,484)
15. (385,5)
16. (196,2,1421)
17. (385,5,66)
18. (525,3923)
19. (135.6,I,21-22)
20. (495,9,52)
21. (158,3,124,161; 460,5, Tablice 21,22,74; 533,6,415)
22. (360,8,109)
23. (230.25,24)
24. (153,4,173)
25. (230.25,4-5,Tabela 1)
26. (101,67)
27. (360,9,14)
28. (259,8309)
29. (360.9,14-15). Zobacz także (531.3,Ryc. 459,460).
30. (405,7,230; 485,2)
31. (360,85,143-144)
32. (125.1,20-21)
33. (360,85,144)
34. (360,9,16)
35. (50,1,211)
36. (379,9,162, 170-171)
37. (360.9,16)
38. (99,2; 360,85151; 473,6155)
39. (360.9,16)
40. (480,3619)
41. (470.2,13)
42. (159,5694)
43. (159,5689)
44. (159,5685)
45. (68,3775)
46. (63,8; 390,2293; 360,9,14; 447,2,77-78)
47. (390,2,293)
48. (250,2418)
49. (250,8,183-184)
50. (329,5,70-71)
51. (250,8,183-188)
52. (360.9,7). Zobacz także (360.8,109; 360.85,29).
53. (90.1,485). Zobacz także (360.3,13,20,21).
54. (360,8,115).
55. (360.3,20).
56. (360.3,20)
57. (360,9,6)
58. (360,7578)
59. (360,7582)
60. (360,7,588). Zobacz także (63,4,165-166; 470,2,Tabela I).
61. (232,5; 360,7)
62. (360,8122)
63. (178,5,199a)
164. (400,5378)
165. (72,5)
166. (328,28)
167. (15.4,63)
168. (400,5,378).
169. (400,5378)
170. Zobacz między innymi (118.5) i (595.4).
171. (36,1; 277,4; 287,1).
172. (400,5383)
173. (380,5)
174. (30,8100; 385,5)
175. (286,3)
176. (595,4,24)
177. (390,8,42)
178. (595.4,26-27)
179. (286,3,29)
180. (193,4)
181. (400,5,382-383)
182. (602,9)
183. (400,5379)